Transkription (Biologie)
| Übergeordnet |
| Genexpression RNA-Biosynthese |
| Untergeordnet |
| Initiation Elongation Termination |
| Gene Ontology |
|---|
| |
Als Transkription (von spätlateinisch transcriptio „Übertragung“ zu lateinisch transcribere „um-/ überschreiben“) wird in der Genetik die Synthese von RNA anhand einer DNA als Vorlage bezeichnet. Die dabei entstehende RNA lässt sich größtenteils in drei Gruppen einteilen: mRNA (messenger RNA) (zur Proteinbiosynthese) sowie tRNA (transfer RNA) und rRNA (ribosomale RNA). Die Transkription ist, wie auch die Translation, ein wesentlicher Teilprozess der Genexpression.

Bei der Transkription wird ein Gen abgelesen und als RNA-Molekül vervielfältigt, das heißt ein spezifischer DNA-Abschnitt dient als Vorlage (Matrize, englisch template) zur Synthese eines neuen RNA-Strangs. Bei diesem Vorgang werden die Nukleinbasen der DNA (A – T – G – C) in die Nukleinbasen der RNA (U – A – C – G) umgeschrieben. Anstelle des Thymins kommt Uracil und anstelle der Desoxyribose kommt Ribose in der RNA vor.
Der Vorgang der Transkription verläuft bei Eukaryoten und Prokaryoten grundsätzlich gleich. Unterschiede gibt es bei der Steuerung und bei der anschließenden Modifikation. Bei Prokaryoten erfolgt die Steuerung über einen Operator, während bei den Eukaryoten die Regulation über einen Enhancer oder Silencer geregelt werden kann, der jeweils dem Promotor vor- oder nachgeschaltet ist. Weiterhin erfolgt bei Prokaryoten die Transkription im Cytoplasma der Zelle, bei Eukaryoten im Zellkern (Karyoplasma). Bei Eukaryoten wird außerdem die prä-mRNA während beziehungsweise nach ihrer Synthese noch prozessiert, bevor sie aus dem Zellkern in das Cytoplasma transportiert wird. Nach der Transkription erfolgt im Cytoplasma am Ribosom die Translation der mRNA in ein Protein.
Synthese der mRNA
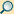
- Synthese der mRNA:
- Allgemeiner Ablauf: Der RNA-Polymerase-Proteinkomplex setzt sich an eine Promotor genannte DNA-Region (siehe dazu den Hauptartikel Initiation). Die RNA-Polymerase entspiralisiert die Doppelhelix und legt so jeweils ca. 10–20 Basen der DNA zur Paarung frei. Am codogenen Strang (Abb.: Antisense-Strang) der DNA lagern sich durch Basenpaarung komplementäre Ribonukleotide an. Sie werden unter Eliminierung von Pyrophosphat aus den Nukleosidtriphosphaten durch eine Esterbindung zwischen Phosphat und Ribose miteinander verknüpft. Die Ableserichtung der DNA verläuft vom 3’-Ende zum 5′-Ende, die Synthese der komplementären RNA dementsprechend von 5’ nach 3’ . Die RNA-Polymerase benötigt keinen Primer, am Terminator wird die Transkription beendet. Danach wird das mRNA-Transkript entlassen und die Polymerase löst sich von der DNA (siehe dazu den Hauptartikel Termination).
- Bei der eukaryotischen mRNA-Synthese kommen zum gerade beschriebenen Ablauf noch die Synthese einer Cap-Struktur am 5’-Ende der mRNA hinzu, die deren Schutz dient und als Signal für den Export aus dem Zellkern fungiert. Dieses sogenannte Capping passiert bereits, wenn das Transkript nur wenige Nukleotide lang ist, also noch vor der Elongation.
- Weitere Verarbeitung der mRNA:
- Bei Prokaryoten gelangt die mRNA nach dem Kopiervorgang direkt zu den Ribosomen, häufig lagern sich auch bereits Ribosomen an die noch entstehende Kette an und beginnen die Translation, bevor die Transkription beendet ist (Poly-Ribosom-Complex).
- Bei Eukaryoten verlässt das RNA-Transkript selbst noch nicht den Zellkern. Die im ersten Teil der Transkription entstandene RNA wird als unreife RNA, prä-mRNA oder hnRNA (heterogene nucleäre RNA) bezeichnet. Sie wird am 3′-Ende durch Polyadenylierung (Tailing) und Splicing noch prozessiert. Durch alternatives Splicing können aus demselben DNA-Abschnitt unterschiedliche mRNA-Moleküle entstehen. Die sogenannte reife mRNA verlässt durch eine Kernpore den Zellkern und gelangt damit ins Zytoplasma, wo sie mit den Ribosomen in Wechselwirkung treten kann.
Synthese der tRNA und der rRNA
Die Transfer-RNA (tRNA) und die ribosomale RNA (rRNA) werden durch zwei andere RNA-Polymerasen an der DNA synthetisiert, die beide nach einem anderen Prinzip als die der RNA-Polymerase II arbeiten. Sie gleichen denen der Prokaryoten, bei denen dieselbe RNA-Polymerase als Katalysator tätig ist. Bei Eukaryoten erfolgt die Synthese der tRNA, der 5S rRNA und der 7SL-RNA durch die RNA-Polymerase III, die Synthese der rRNA und teilweise auch der sn-RNA (small-nuclear RNA) durch die RNA-Polymerase I, die Synthese der m-RNA durch die RNA-Polymerase II.
Ende der Transkription
In eukaryotischen Zellen kann die RNA-Polymerase das Ende eines Gens nicht von alleine erkennen, sie braucht dazu Hilfsfaktoren, die mit der Polymerase in Wechselwirkung treten. Diese Proteinkomplexe erkennen die Polyadenylierungsstelle (5’-AAUAAA-3’), schneiden die RNA und leiten die Polyadenylierung ein, während die RNA-Polymerase gleichzeitig weiterarbeitet. Ein Modell für die Termination der Transkription ist, dass das noch immer weiter wachsende, nutzlose RNA-Ende von einer Exonuklease (Rat1) abgebaut wird, und zwar schneller, als es von der Polymerase verlängert wird. Erreicht die Exonuklease die Transkriptionsstelle, löst sich die Polymerase von der DNA, die Transkription ist endgültig beendet (Torpedo model of transcriptional termination). Darüber hinaus scheinen weitere Proteinkomplexe (z.B.: TREX) für eine Termination wichtig zu sein.
Archaeelle Transkription
Die Gene der Archaea besitzen im Promotor eine TATA-Box genannte Konsensussequenz. Am Promotor binden die zwei Initiationsfaktoren der Archaea, TBP und TFB. An diese bindet wiederum eine Polymerase, die ortholog zur eukaryotischen RNA-Polymerase II ist und aus zwölf Untereinheiten besteht.
Die Transkription ist in vielen Fällen an das darauf folgende Spleißen der mRNA gekoppelt, bevor die mRNA aus einer Kernpore schwimmt oder nun direkt schon weiter zu einem Protein synthetisiert wird.
Bakterielle Transkription
Im Gegensatz zu den Eukaryoten besitzen Bakterien nur eine RNA-Polymerase. Das Core- beziehungsweise Minimal-Enzym besteht aus vier Untereinheiten (2× α, β, β’), das die Transkription katalysiert, sie aber nicht initiiert. Der Core des Enzyms wirkt wechselseitig mit der losen Sigma-Untereinheit und es bildet sich das Holo-Enzym (2× α, β, β’, σ (Sigma)), welches die Initiation durchführen kann (Sigma ermöglicht das Entlanggleiten an der DNA und Auffinden der Pribnow-Box des Promotors). Die Sigma-Untereinheit bindet am Promotor des Nicht-Matrizenstranges und löst dort die Wasserstoffbrücken zwischen den Basenpaaren; sie besitzt eine Helicasefunktion, was die wichtigste Funktion dieser Polymerase ist.
Funktion der α-Untereinheit ist zum einen durch die aminoterminale Domäne bedingt der Erhalt und die Stabilität der Struktur, zum anderen durch die carboxyterminale Domäne eine Bindung an den Promotor und die Wechselwirkung mit Transkriptions-regulatorischen Elementen.
Die β- und β’-Untereinheiten wirken zusammen und sorgen für die Bindung an die DNA-Matrize und für eine wachsende RNA-Kette.
Die σ-(Sigma)-Untereinheit erkennt Transkriptionsstartpunkte. Es gibt mehrere Sigma-Untereinheiten, die verschiedene Gengruppen erkennen. Am weitesten verbreitet ist die σ70 Untereinheit.
Die ω-(Omega)-Untereinheit dient der Stabilisierung und der Strukturaufrechterhaltung und ist nicht zwingend notwendig.
Reverse Transkription
In einigen Viren gibt es den Sonderfall der reversen Transkription, bei der nach einer RNA-Vorlage DNA hergestellt wird.
Literatur
- Rolf Knippers: Molekulare Genetik. 9. komplett überarbeitete Auflage. Thieme, Stuttgart u. a. 2006, ISBN 3-13-477009-1.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 18.09. 2025