Virophagen

Als Virophagen[A. 1] klassifiziert man nicht-taxonomisch einen speziellen Typ vergleichsweise großer Satellitenviren aus dem Phylum Preplasmiviricota, die sich nur in Co-Infektion mit Riesenviren des Phylums Nucleocytoviricota (NCLDVs) als Helferviren in eukaryotischen Wirtszellen replizieren können. Virophagen replizieren (anders als die Begriffliche Anlehnung an Bakteriophagen nahelegen könnte) nicht „in“ anderen Viren, sondern nutzen als Satellitenviren den Syntheseapparat der Helferviren – und treten dabei in eine Konkurrenz zu ihnen.[1][2] Dabei werden Genprodukte des Helfervirus genutzt, die sonst nur bei eukaryotischen Zellen (Eucyten) die Proteinsynthese ermöglichen, daher werden die Helferviren der Virophagen auch Mamaviren[A. 2] genannt. In der Regel wird die Proteinsynthese des Mamavirus dabei mehr oder weniger beeinträchtigt.[3][4] Virophagen sind daher (bzgl. der Mamaviren) parasitäre Satellitenviren. Daher werden die Helfer- oder Mamaviren auch als „virale Wirte“, die co-infizierten Eucyten zur Abgrenzung dann auch als „zelluläre Wirte“ bezeichnet. Die Virophagen der Riesenviren sind im Vergleich zu Satellitenviren anderer Helferviren ebenfalls vergleichsweise riesig und haben auch ein komplexeres Genom als diese.[5]
Virophagen sind sowohl bzgl. ihrer Kapsidgröße als auch bzgl. ihres Genoms viel kleiner als ihre Riesenviren-Wirte, aber andererseits viel größer als gewöhnliche Satellitenviren, deren Helferviren ja ebenfalls kleiner als die Riesenviren der NCLDV sind.
Forschungsgeschichte und prominente Beispiele
Der erste Virophage namens Sputnik wurde im Jahr 2008 von Bernard La Scola und Didier Raoult von der Université de la Méditerranée in Marseille in den Rohren eines Kühlwassersystems in Paris entdeckt. Er parasitiert das Acanthamoeba castellanii mamavirus (AcMV, ein Stamm der Spezies Mimivirus bradfordmassiliense).[4]
Die bislang (Stand 3. März 2025) vom International Committee on Taxonomy of Viruses (ICTV) offiziell bestätigten Virophagen gehören fast alle zu einer Klasse, den Virophaviricetes (ehemals Maveriviricetes, hervorgegangen aus der früheren Familie Lavidaviridae), mit folgenden Gattungen:[7][8]
- Ordnung Divpevirales, Familie Ruviroviridae
- Ordnung Lavidavirales, Familie Maviroviridae
- Mavirus mit einer einzigen Spezies Mavirus cafeteriae
– diese parasitieren Mamaviren der Aliimimivirinae (Mimiviridae der Gruppe II).
- Mavirus mit einer einzigen Spezies Mavirus cafeteriae
- Ordnung Mividavirales, Familie Sputniviroviridae
- Sputnikvirus mit den beiden Spezies
Sputnikvirus mimiviri und Sputnikvirus zamilonensen
– diese parasitieren Mamaviren der Megamimivirinae (Mimiviridae-Gruppe I)
- Sputnikvirus mit den beiden Spezies
Sputnikvirus mimiviri und Sputnikvirus zamilonensen
- Ordnung Priklausovirales
- Familie Burtonviroviridae: Burquivirus mit der einzigen Spezies Burquivirus flavolapense (Yellowstone Lake virophage 5, YSLV5).[12]
- Familie Dishuiviroviridae: Essdubovirus mit der einzigen Spezies Essdubovirus chlorellae (Chlorella virophage isolate SW01 aus dem Dishui Lake, China)[13]
- Familie Gulliviroviridae: Invirovirus mit der einzigen Spezies Invirovirus crochense (Lake Croche Virophage IMG_VR_1276)[14]
- Familie Omnilimnoviroviridae: Panaquavirovirus mit der einzigen Spezies Panaquavirovirus qinghaense (Qinghai Lake virophage, QLV, Fundort von QLV ist der Qinghai-See)[15][16]
Abweichend zur Klasse Aquintoviricetes gehört Tetrivirus crimaeaense mit dem Tetraselmis-viridis-Virus S1 (TvV-S1).
_Guarani_Mimivirus.png)
_Guarani_progeny.png)
Es gibt zudem – meist aus der Metagenomik – viele weitere Hinweise und Vorschläge zu Virophagen innerhalb der Virophaviricetes (alias Maveriviricetes bzw. Lavidaviridae in älteren Arbeiten) oder auch außerhalb; einen Überblick über die Virophagen geben beispielsweise Zhou et al. (2013),[17] Gong et al. (2016),[18] und Chen et al. (2018):[19]
Einige prominente Beispiele sind:
- der „Organic-Lake-Virophage“ (engl. „Organic Lake virophage“, OLV),(N,G)[A. 3][2]
- der „Guarani virophage“[A. 4][20][21][22]
- die „Dishui-Lake-Virophagen“ 1 bis 8 (DSLV1 bis DSLV8)[A. 5][18][23]
- die „Yellowstone-Lake-Virophagen“ 1 bis 4, 6 und 7 (YSLV1 bis YSLV4, YSLV6, YSLV7)[A. 6][24][25]
- die TBE-Virophages (TBE: englisch Trout Bog epilimnion) und TBH-Virophagen (TBH: engl. Trout Bog hypolimnion)[A. 7][9]
- Loki’s Castle virophages (LC-Virophagen)[A. 8][26]
- „Preplasmiviricota sp. Gezel-14T“ (PLV ‘Gezel-14T) alias Phaeocystis globosa Virus Virophage (PgVV)[A. 9][27][28][29][6][5]
Gezel-14T gehört möglicherweise wie TvV-S1 nicht zu den Virophaviricetes (alias Maveriviricetes), sondern in einen anderen Zweig der Preplasmiviricota (bzw. dem die Virophaviricetes enthaltenden Subphylum Polisuviricotina).
Weitere Gensequenzen von Virophagen hat man unter anderem auch an folgenden Stellen gefunden:
- Punta Cormorant, eine salzige Lagune auf Floreana (Santa María, englisch Charles, Galápagos-Inseln)[2][30][1]
- Einem Ocean Upwelling auf Fernandina (englisch Narborough Island, ebenfalls Galápagos-Inseln)[2][30][1]
- Delaware Bay Ästuar (New Jersey, Ostküste der USA)[2][30][1]
- Gatúnsee (englisch Lake Gatun, ein Süßwassersee in Panama, Mittelamerika)
- Gossenköllesee (Österreich)[31]
Eine Liste nicht-klassifizierter Virophagen findet sich in der Taxonomie das National Center for Biotechnology Information (NCBI).[32]
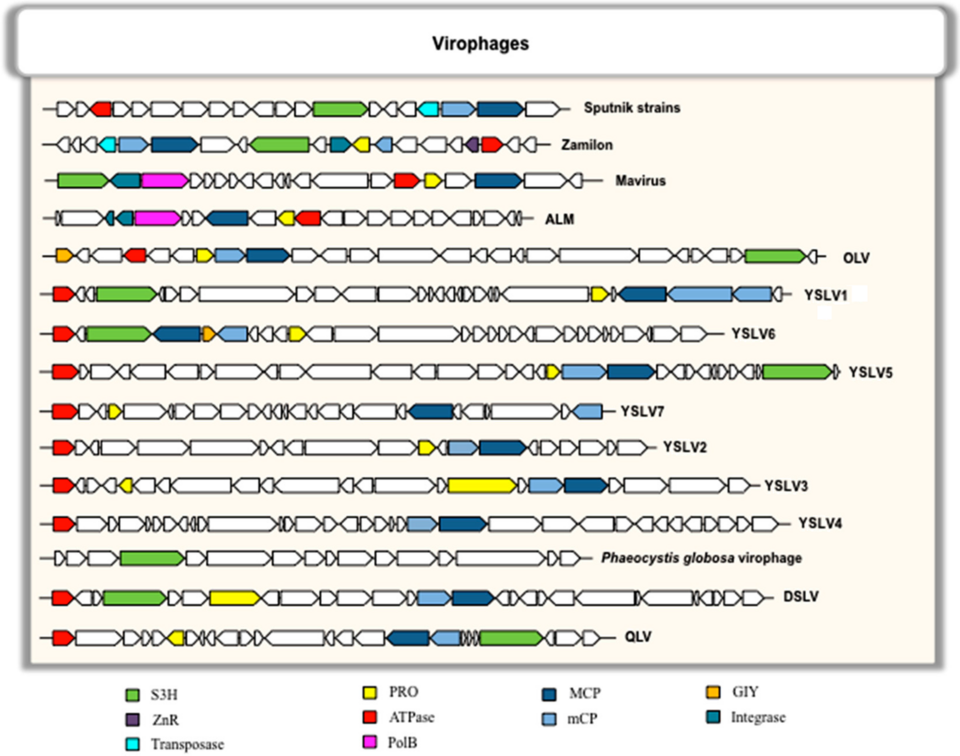
Genom
Replikationszyklus
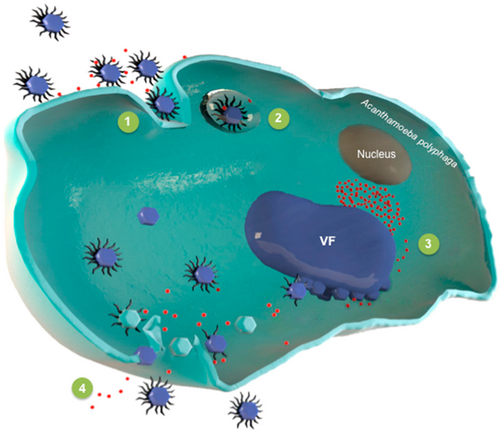
Der Replikationszyklus der Virophagen[A. 10] unterscheidet sich grundlegend von dem gewöhnlicher Satellitenviren.
Normalerweise initiiert ein Satellitenvirus die Expression und Replikation seines Genoms im Zellkern der Wirtszelle mit Hilfe seiner dortigen Maschinerie und geht dann in das Zytoplasma. Dort findet es die Morphogenese-Maschinerie seines Helfervirus, mit der dieses seine Virionen (Virusteilchen) zusammenbaut (Assembly). Diese wird vom Satellitenvirus gekapert, um seine eigenen Nachkommen zu produzieren.[6]
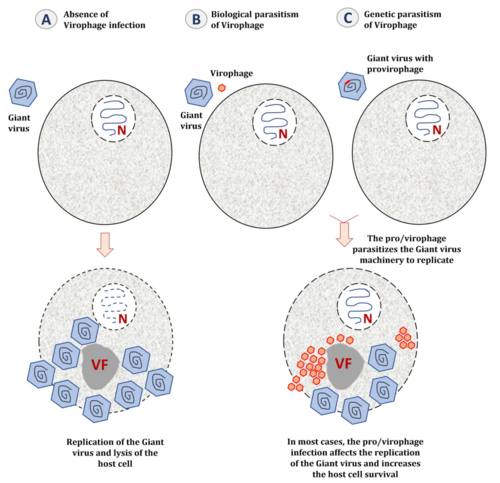
Bei den Virophagen der Familie Lavidaviridae wird angenommen, dass die Replikation der Virophagen vollständig in der Virusfabrik des Riesenvirus-Wirts stattfindet und daher vom Expressions- und Replikationskomplex des Riesenvirus abhängt.[6] Wenn die Wirtszelle nur vom Riesenvirus infiziert wird, baut dieses im Zytoplasma eine Virusfabrik (VF) auf, um sich zu replizieren und neue Virionen zu erzeugen. Die Wirtszelle wird am Ende seines Replikationszyklus gewöhnlich lysiert, d. h. unter Freisetzung der Virus-Nachkommenschaft zum Platzen gebracht und so zerstört.[6]
Wenn die Wirtszelle mit einem Riesenvirus und einem Virophagen koinfiziert ist, parasitiert letzterer in der Virusfabrik des Riesenvirus. Die Anwesenheit von Virophagen kann die Infektiosität des Riesenvirus daher ernsthaft beeinträchtigen, indem sie seine Replikationseffizienz verringert und damit indirekt das Überleben der Wirtszelle erhöht.[6]
Virophaviricetes
Die meisten derzeit (Stand März 2025) bekannten Virophagen gehören der Klasse Virophaviricetes an, einige der Klasse Aquintoviricetes. Sie gehören dem Subphylum Polisuviricotina des Phylums Preplasmiviricota an.[7]
Provirophagen
Ein Sonderfall ist, wenn sich der Virophage als „Provirophage“ in das Genom des Mamavirus integriert. Sowohl die Virophagen (als Mitglieder der Preplasmiviricota), als auch die Mamaviren (als Riesenviren Mitglieder der Nucleocytoviricota) sind dsDNA-Viren. Der Provirophage wird während der Replikation des Riesenvirus exprimiert. Der Virophage wird dann in der Virusfabrik des Riesenvirus produziert und hemmt die Replikation des Riesenvirus, wodurch wieder das Überleben der Wirtszelle erhöht wird – Abbildung oben, Teil (C). Ein Beispiel ist Sputnik 2, der sich in das Genom seines viralen Wirts, einem Mimivirus, integrieren kann.[6]
Defektive interferierende Viren
In der Funktionsweise ähnlich sind DIVs („Defektive interferierende Viren“, englisch defective interfering viruses, auch defective interfering particles, DIPs). Dies sind natürlich entstandene oder künstlich erzeugte defektive Abarten eines Virus-Wildtyps, die den Wildtyp als Helfervirus benötigen und wie Parasiten dessen Replikation stören können.[33]
Transpovirons
Eine weitere verwandte Klasse mobiler genetischer Elemente sind die Transpovirons, zuerst entdeckt im Genom des „Lentille-Virus“ („Mimivirus lentille“, auch „Acanthamoeba polyphaga lentillevirus“).[34][6][35]
Phylogenie
Eugene V. Koonin und Mart Krupovic schlugen 2017 auf Grundlage von Sequenzen der DNA-Polymerase für die Phylogenie Polinton-ähnlicher mobiler genetischer Elemente (MGEs) und ihrer Abkömmlinge (Viren der Preplasmiviricota) folgendes Szenario vor (ergänzt um moderne Bezeichnungen nach ICTV MSL#40v1, 3. März 2025):[35][7]
| |||||||||||||||||||||||||||||||||||||
Moderne Taxonomien ordnen die Bidnaviridae jedoch dem Realm Monodnaviria zu, während die anderen oben genannten Virengruppen dem Realm Varidnaviria angehören.[7]
Weblinks
- Christelle Desnues, Bernard La Scola, Natalya Yutin, Ghislain Fournous, Catherine Robert, Saïd Azza, Priscilla Jardot, Sonia Monteil, Angélique Campocasso, Eugene V. Koonin, Didier Raoult: Provirophages and transpovirons as the diverse mobilome of
iant viruses. In: PNAS. Band 109, Nr. 44, 30. Oktober 2012, S. 18078–18083;
 doi:10.1073/pnas.1208835109 (englisch).
doi:10.1073/pnas.1208835109 (englisch). - Graziele Oliveira, Bernard La Scola, Jônatas Abrahão: Giant virus vs amoeba: fight for supremacy. In: Virology Journal. Band 16, Nr. 1, Artikel 126, veröffentlicht: 4. November 2019;
doi:10.1186/s12985-019-1244-3
( Volltext, PDF auf: researchgate.net, englisch).
- Laurie O’Keefe: Sizing Up Viruses. In: The Scientist. ( Volltext als PDF Auf: cdn.the-scientist.com)
Illustration zu Didier Raoult: Viruses Reconsidered. In: The Scientist. 28. Februar 2014: Grafische Darstellung der Größenverhältnisse
( Volltext, auf: the-scientist.com, englisch).
- David Paez-Espino, Jinglie Zhou, Simon Roux, et al.: Diversity, evolution, and classification of virophages uncovered through global metagenomics. In: Microbiome. Band 7, Nr. 157, 10. Dezember 2019;
doi:10.1186/s40168-019-0768-5 (englisch).
Anmerkungen
- ↑ Singular: Virophage, der; von lat. virus, -i, n. „Gift, Saft, Schleim“ und altgriechisch φαγεῖν phageín, „fressen“, Virophage bedeutst also „Viren-Esser“
- ↑ Das erste so bezeichnete Mamavirus ist das Acanthamoeba castellanii mamavirus (AcMV), ein Stamm der Spezies Mimivirus bradfordmassiliense.
- ↑ OLV parasitiert „Organic Lake Phycodnavirus“ (OLPV, Mesomimiviridae), Fundort: Organic Lake.
- ↑ Fundort des Guarani-Virophagen ist der Pampulha-See, Belo Horizonte.
- ↑ Die DSL-Virophagen parasitieren offenbar Phycodnaviridae (Dishui Lake phycodnaviruses, DSLPVs) und Mimiviridae (sic!, Dishui Lake large alga viruses, DSLLAVs, daher ist Mimiwiridae wohl s. l. zu verstehen und es handelt sich vermutlich um Algen-infizierende Imitervirales), Fundort Dishui Lake
- ↑ Die YSLVs sind zu unterscheiden von ihren mutmaßlichen Wirten „Yellowstone Lake Phycodnavirus“ (YSLPV) alias „Yellowstone Lake Mimivirus“ oder „Yellowstone Lake Giant Virus“ (YSLGV), Fundort: Yellowstone Lake.
- ↑ Die TBE- bzw. TBH-Virophagen wurden in den Oberflächenschichten (Epilimnion) respektive in den Tiefen (Hypolimnion) des Moorsees Trout Bog Lake, Wisconsin, gefunden.
- ↑ Die mutmaßlichen LC-Virophagen stammen vom Genite des Schwarzen Rauchers Lokis Schloss.
- ↑ Gezel-14T parasitiert Phaeocystis-globosa-Virus PgV-14 und 16 (PgV-14T und PgV-16T, beide Spezies Tethysvirus hollandense).
- ↑ Analysiert sind derzeit (2. Oktober 2023) nur Lavidaviridae, es dürfte aber ähnlich für alle Virophagen der Preplasmiviricota gelten.
Einzelnachweise
- ↑ Hochspringen nach: a b c
d Jan Osterkamp: Freundliche Feinde des Feindes.
Auf: spektrum.de – News vom 31. März 2011; zuletzt abgerufen 19. Oktober 2025.
- ↑ Hochspringen nach: a b c d
e Sheree Yau, Federico M. Lauro, Matthew Z. DeMaere, Mark V. Brown et al.: Virophage control of antarctic algal host–virus dynamics. In:
Proceedings of the National Academy of Sciences. Band 108, Nr. 15, 12. April 2011 (online 28. März 2011), S. 6163–6168;
doi:10.1073/pnas.1018221108,
PMC 3076838 (freier Volltext),
PMID 21444812 (englisch).
- ↑
Ulrich Kuhnt: Sind Viren Lebewesen?. Max-Planck-Institut für biophysikalische Chemie: Archiv Biologie. Memento im Webarchiv vom 21. Januar 2018.
- ↑ Hochspringen nach: a b La Scola et al.: The virophage as a unique parasite of the giant mimivirus.
In: Nature. Band 455, 4. September 2008, S. 100–105;
doi:10.1038/nature07218 (englisch).
- ↑ Hochspringen nach: a b c d
Meriem Bekliz, Philippe Colson, Bernard La Scola: The Expanding Family of Virophages. In: MDPI Viruses. Band 8, Nr. 11 (Special Issue Viruses of Protozoa), 23. November 2016, S. 317;
doi:10.3390/v8110317 (englisch).
- ↑ Hochspringen nach: a b c d
e f g h
i j Said Mougari, Dehia Sahmi-Bounsiar, Anthony Levasseur, Philippe Colson, Bernard La Scola: Virophages of
Giant Viruses: An Update at Eleven. In: Viruses. Band 11,
Nr. 8, 2019,
ISSN 1999-4915, S. 733,
doi: 10.3390/v11080733,
PMID 31398856,
PMC 6723459 (freier Volltext) – (englisch).
- ↑ Hochspringen nach: a b c d
ICTV: MSL #40.v1, 3. März 2025. (auto. Download)
- ↑ ICTV: VMR zu MSL #40.v1, 3. März 2025. (auto. Download)
- ↑ Hochspringen nach:a b Simon Roux, Leong-Keat Chan, Rob Egan, Rex R. Malmstrom, Katherine D. McMahon, Matthew B. Sullivan:
Ecogenomics of virophages and their giant virus hosts assessed through time series metagenomics. In: Nature Communications.
Band 8, Nr. 858, 11. Oktober 2017,
doi: 10.1038/s41467-017-01086-2,
PMID 29021524,
PMC 5636890 (freier Volltext) – (englisch). –
( OSTI: DOE Pages Auf: osti.gov Office of Scientific and Technical Information Energieministerium der Vereinigten Staaten,
Volltext als PDF Auf: nature.com); siehe insbes. Fig. 1 und Fig. 2. Anm.: In Fig. 2 muss es wie in Fig. 1 YSLV statt YLSV
heißen (Yelloxstone Lake Virophage).
- ↑ Natalya Yutin, Vladimir V. Kapitonov, Eugene V. Koonin: A new family of hybrid virophages from an animal gut
metagenome. In: Biology Direct. Band 10,
Nr. 19, 25. April 2015,
ISSN 1745-6150,
doi: 10.1186/s13062-015-0054-9,
PMID 25909276,
PMC 4409740 (freier Volltext) – (englisch).
- ↑ NCBI Nucleotide: Gut metagenome contig-21000060, … Accession: AUXO017923253.
- ↑ NCBI Nucleotide: MAG: Yellowstone Lake virophage 5, complete genome. Accession: KM502589.
- ↑ NCBI Nucleotide. Chlorella virophage isolate SW01, complete genome. Accession: OL828819.
- ↑ NCBI Nucleotide: MAG: Lavidaviridae sp. isolate IMG_VR_1276, complete genome. Accession: OQ836152.
- ↑ Seungdae Oh, Dongwan Yoo, Wen-Tso Liu: Metagenomics Reveals a Novel Virophage Population in a Tibetan Mountain Lake. In:
Microbes and Environments. Band 31,
Nr. 2, Juni 2016,
ISSN 1342-6311, S. 173–177,
doi: 10.1264/jsme2.ME16003,
PMID 27151658,
PMC 4912154 (freier Volltext) – (englisch).
- ↑ NCBI Nucleotide: MAG: Qinghai Lake virophage, complete genome. Accession: KJ854379.
- ↑ Jinglie Zhou, Weijia Zhang, Shuling Yan et al.: Diversity of Virophages in Metagenomic Data Sets. In:
Journal of Virology. Band 87, Nr. 8, Februar 2013,
S. 4225–4236,
doi: 10.1128/JVI.03398-12,
PMID 23408616 (englisch).
- ↑ Hochspringen nach: a b Chaowen Gong, Weijia Zhang, Xuewen Zhou, Hongming Wang et al.:
Novel Virophages Discovered in a Freshwater Lake in China. In: Frontiers in Microbiology. Band 7, 22. Januar 2016,
ISSN 1664-302X,
doi: 10.3389/fmicb.2016.00005,
PMID 26834726 (englisch).
- ↑ Hao Chen, Weijia Zhang, Xiefei Li, Yingjie Pan, Shuling Yan, Yongjie Wang: The genome of a prasinoviruses-related freshwater
virus reveals unusual diversity of phycodnaviruses. In: BMC Genomics. Band 19,
Nr. 49, 15. Januar 2018,
ISSN 1471-2164,
doi: 10.1186/s12864-018-4432-4,
PMID 29334892,
PMC 5769502 (freier Volltext) – (englisch). – Anmerkung: OLV ist hier Abkürzung für Organic Lake Virophage, OlV für
Ostreococcus lucimarinus virus (Gattung Prasinovirus).
- ↑ Clara Rolland, Julien Andreani, Amina Cherif Louazani, Sarah Aherfi, Rania Francis, Rodrigo Rodrigues, Ludmila Santos Silva, Dehia Sahmi, Said Mougari,
Nisrine Chelkha, Meriem Bekliz, Lorena Silva, Felipe Assis, Fábio Dornas, Jacques Yaacoub Bou Khalil, Isabelle Pagnier, Christelle Desnues, Anthony Levasseur, Philippe Colson, Jônatas Abrahão, Bernard La Scola:
Discovery and Further Studies on Giant Viruses at the IHU Mediterranee Infection That Modified the Perception of the Virosphere. In: MDPI Viruses.
Band 11, 4 (März/April), 2019,
ISSN 1999-4915, S. 312,
doi: 10.3390/v11040312,
PMID 30935049,
PMC 6520786 (freier Volltext).
- ↑ Said Mougari, Meriem Bekliz, Jonatas Abrahao, Fabrizio Di Pinto, Anthony Levasseur, Bernard La Scola: Guarani Virophage,
a New Sputnik-Like Isolate From a Brazilian Lake. In: Frontiers in Microbiology. Band 10, 3. Mai 2019,
ISSN 1664-302X,
doi: 10.3389/fmicb.2019.01003,
PMID 31130943,
PMC 6510173 (freier Volltext). –
( Volltext als PDF Auf: semanticscholar.orgPDF)
- ↑ NCBI BioSample: Guarani virophage genome.
- ↑ Shengzhong Xu, Liang Zhou, Xiaosha Liang, Yifan Zhou, Hao Chen, Shuling Yan, Yongjie Wang: Novel Cell-Virus-Virophage Tripartite Infection Systems Discovered in the
Freshwater Lake Dishui Lake in Shanghai, China. In: ASM Jounals: Journal of Virology, Band 94, Nr. 11, 18. Mai 2020, e00149-20;
doi:10.1128/JVI.00149-20,
PMID 32188734,
PMC 7269436 (freier Volltext) (englisch).
- ↑ Weijia Zhang, Jinglie Zhou, Taigang Liu, Yongxin Yu et al.: Four novel algal virus genomes discovered from Yellowstone
Lake metagenomes. In: Scientific Reports. Band 5, Nr. 1, 2015,
ISSN 2045-2322, S. 15131,
doi: 10.1038/srep15131,
PMID 26459929 (englisch, Figur 6).
- ↑ Virus-Host DB: Yellowstone lake mimivirus. Auf:
genome.jp; zuletzt abgerufen am 19. Oktober 2025.
- ↑ Disa Bäckström, Natalya Yutin, Steffen L. Jørgensen, Jennah Dharamshi, Felix Homa, Katarzyna Zaremba-Niedwiedzka, Anja Spang, Yuri I. Wolf,
Eugene V. Koonin, Thijs J. G. Ettema: Virus Genomes from Deep Sea Sediments Expand the Ocean Megavirome and Support Independent Origins of Viral Gigantism. In:
mBio. Band 10, Nr. 2, 2019,
ISSN 2150-7511,
doi: 10.1128/mBio.02497-18,
PMID 30837339 (englisch).
- ↑ NCBI Taxonomy Browser:
Phaeocystis globosa virus virophage, syn. Preplasmiviricota sp. Gezel-14T (species), Nucleotide:
txid1335638[Organism:noexp], Preplasmiviricota sp. Gezel-14T.
- ↑ NCBI Nucleotide:
txid1335638[Organism:noexp] AND PgV-16T.
- ↑ Sheila Roitman, Andrey Rozenberg, Tali Lavy, Corina P. D. Brussaard, Oded Kleifeld, Oded Béjà: Isolation and infection cycle of a polinton-like virus virophage
in an abundant marine alga. In: Nature Microbiology. Band 8, 26. Januar 2023, S. 332–346;
doi: 10.1038/s41564-022-01305-7,
PMID 36702941 (englisch).
- ↑ Hochspringen nach: a b c Meriem Bekliz, Jonathan Verneau, Samia Benamar
et al.: A New Zamilon-like Virophage Partial Genome Assembled from a Bioreactor Metagenome. In: Frontiers in Microbiology.
Band 6, 27. November 2015,
ISSN 1664-302X,
doi: 10.3389/fmicb.2015.01308,
PMID 26640459 (englisch).
- ↑ Christopher M. Bellas, Ruben Sommaruga: Polinton-like viruses are abundant in aquatic ecosystems. In: BMC: Microbiome. Band 9, Nr. 13, 12. Januar 2021;
doi:10.1186/s40168-020-00956-0.
- ↑ NCBI Taxonomy Browser: unclassified virophages.
- ↑ Fighting COVID With COVID: Driving the Disease to Extinction With a Defective Version of the
SARS-CoV-2 Virus. Auf: scitechdaily.com vom 20. August 2021.
- ↑ Natalya Yutin, Didier Raoult, Eugene V. Koonin: Virophages, polintons, and transpovirons: a complex evolutionary network
of diverse selfish genetic elements with different reproduction strategies. In: Virology Journal. 10. Jahrgang, Nr. 1, 2013,
S. 158,
doi: 10.1186/1743-422X-10-158,
PMID 23701946,
PMC 3671162 (freier Volltext) – (englisch).
- ↑ Hochspringen nach: a b Eugene V. Koonin, Mart Krupovic: Polintons,
virophages and transpovirons: a tangled web linking viruses, transposons and immunity. In: Current Opinion in Virology, Band 25, August 2017, S. 7–15;
doi:10.1016/j.coviro.2017.06.008,
PMID 28672161,
PMC 5610638 (freier Volltext), Epub 30. Juni 2017 (englisch). Siehe insbes.
Gig. 3.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 19.10. 2025